Читайте также:
|
Для радиации характерен кластерный эффект, когда участки ионизации (повреждений) распределяются вдоль молекулы ДНК в компактные кластеры. Понятно, что такой эффект более присущ для плотноионизирующего излучения, чем для редкоионизирующего (рис. 2.5.1) [AR14], но и для последнего имеются значительные отличия в эффекте по сравнению со спонтанными повреждениями ДНК свободными радикалами [AU12].
Еще в начале 1980 гг. было постулировано, что радиация, помимо двунитевых разрывов, может индуцировать возникновение нескольких близко расположенных в пределах короткого фрагмента ДНК повреждений («множественных сайтов повреждений») — кластеров из модифицированных оснований, из находящихся рядом однонитевых разрывов и поврежденного основания (оснований), двунитевого и однонитевого разрывов и др. Была постулирована трудность репарации таких комплексных повреждений, независимо от их химической структуры, по сравнению с отдельными повреждениями [AG6, AW2, AW3, AW4]. Например, попытка репарировать кластер, состоящий из двух модифицированных оснований, расположенных близко на противоположных нитях ДНК, должна приводить к образованию двунитевого разрыва, поскольку одним из этапов репарации оснований является нуклеазный надрез цепи ДНК [AG6]. И действительно, было показано, что в процессе воссоединения пострадиационных разрывов в ДНК уже после облучения могут возникать новые (de novo), не обусловленные радиационным фактором двунитевые разрывы [AS54].
Имеются данные, что исходный уровень двунитевых разрывов, индуцированных в клетке сразу после облучения, слабо коррелирует с последующим биологическим эффектом [AI2]. С этим эффектом коррелируют оставшиеся нерепарированными двунитевые разрывы [AD15], хотя количество таковых и может быть очень мало. Таким образом, среди общего пула двунитевых разрывов выделяются различные классы, отличающиеся по способности к репарации [AD15].
Выводом из двух указанных постулатов явилась гипотеза, согласно которой именно кластеры повреждений («комплексные повреждения») ответственны за летальные и/или мутагенные (канцерогенные) эффекты ионизирующей радиации [AG6, AW2, AW3, AW4].
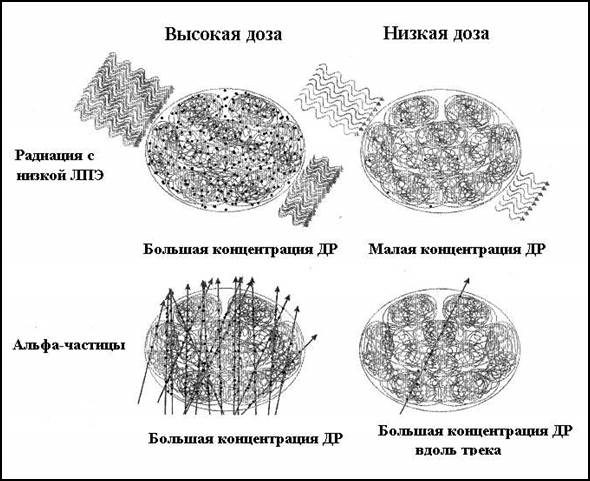
Рисунок 2.5.1. Схема пространственного распределения двунитевых разрывов (ДР) ДНК в ядре после поражения редкоионизирующей радиацией и α-частицами. Низкая доза для последних соответствует здесь малой по определению (пролет одной частицы). По (K. Rothkamm and M. Lobrich, 2002) [AR14].
Вероятность существования сложных комплексных повреждений получила подкрепление в начале 1990-х гг. в модельных экспериментах, имитирующих взаимодействие трека ионизирующей частицы с клеточной мишенью. На модели «Монте-Карло» (исследование пути частиц излучений в воде) было показано, что вдоль треков частиц и с высокой, и с низкой ЛПЭ образуются локальные кластеры ионизации, обусловленные не только прямым поглощением энергии излучения, но и эффектом вторичных низкоэнергетических электронов, «выбиваемых» заряженными частицами. Исходя из параметров поглощения энергии и длины пробега частиц, предсказали реальность кластерного эффекта радиации для ДНК в растворе и в клетке (см. [AG6]), причем оказалось, что с повышением ЛПЭ излучения возрастает не только число, но и сложность кластеров [AN19].
Чем выше ЛПЭ излучения, и, соответственно, больше компактность треков, тем более распределение повреждений в молекуле ДНК отличается от случайного, и с тем большей вероятностью возникают комплексные (кластерные) повреждения (см. рис. 2.5.1).
Таким образом, физические модели убеждали в существовании кластерного эффекта излучения. Но только в начале 2000-х гг. были получены экспериментальные данные о наличии в живых клетках комплексных лучевых повреждений (отличных от двунитевых разрывов). Группой исследователей из США во главе с доктором Бетси Сазерланд (B. Sutherland) был разработан метод определения комплексных повреждений ДНК.
Методика основана на том, что некоторые эндонуклеазы E. coli распознают конкретные кластеры: белок Nth — кластеры оксипиримидинов, белок Fpg — оксипуринов, а белок Nfo — кластеры потерянных оснований. Некоторые типы последних кластеров имеют специфическую устойчивость вследствие плотной структуры и для их расщепления используется путресцин. Расщепление ДНК по этим специфическим сайтам приводит к образованию двунитевых разрывов, которые можно детектировать количественно [AG2, AS54, AS57].
На рис. 2.5.2 представлена схема подобного определения. Препарат ДНК или ДНК в составе клетки подвергают воздействию радиации либо оставляют интактной (контроль). Излучение индуцирует как индивидуальные повреждения (однонитевые разрывы, модифицированные основания), так и кластеры повреждений (двунитевой разрыв; модифицированное основание + однонитевой разрыв; несколько модифицированных оснований или участков с их потерей и пр.). Затем следует расщепление кластеров эндонуклеазами Nth, Nfo, Fpg, что приводит к формированию новых двунитевых разрывов, помимо радиационно-индуцированных. На конечном этапе проводят электрофорез препаратов ДНК в нейтральном геле агарозы (при этих условиях двунитевая ДНК не денатурирует).
В пробах, полученных из необлученного материала (треки 1 и 2), белки E. coli распознают только немногие отдельные кластеры, и только малое количество ДНК представлено фрагментами с меньшей молекулярной массой (темные полоски). Индуцированные радиацией двунитевые разрывы уменьшают размер молекулы ДНК (трек 3), а обработка эндонуклеазами, распознающими кластеры, приводит к формированию еще меньших молекул (трек 4). Частоту возникших двунитевых разрывов (первичных и вторичных) количественно определяют путем анализа функции распределения молекулярной массы фрагментов ДНК. Из суммарного числа кластеров вычитают количество «чистых» двунитевых разрывов (то есть, индуцированных исключительно излучением, а не обработкой ДНК эндонуклеазами E. coli).
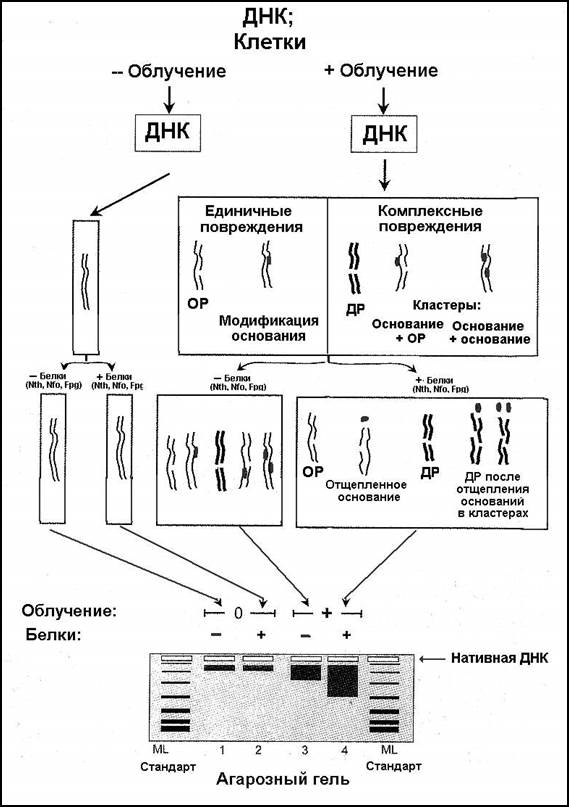
Рисунок 2.5.2. Схема методики определения комплексных повреждений ДНК после облучения, основанная на распознавании кластеров специфическими эндонуклеазами E. coli (подробности в тексте). ОР — однонитевой разрыв, ДР — двунитевой разрыв [AS54, AS57].
Был исследован выход комплексных повреждений ДНК после различных воздействий на двух биологических объектах — очищенной ДНК фага Т7 и культуре моноцитарных клеток человека (линия 28SC). Суммирование результатов позволяет сформулировать следующее [AG2, AS51, AS53, AS54, AS55, AS56, AS57]:
1) Комплексные повреждения ДНК обнаружены после воздействия и ионизирующего, и УФ-излучения, и радиомиметиков.
2) Радиация с высокой ЛПЭ обладает более выраженным кластерным эффектом, чем рентгеновское и g-излучение. УФ-излучение слабо индуцирует кластерные повреждения.
3) Выход четырех типов кластерных повреждений, включая двунитевые разрывы (см. рис. 2.5.2), линейно зависит от дозы рентгеновского и g-излучения в диапазоне 1–100 Гр (137Cs) для ДНК фага Т7 и в диапазоне 50–500 мГр рентгеновского излучения для моноцитов человека (рис. 2.5.3 и 2.5.4 ниже). Авторы приходят к выводу, что для индукции комплексных повреждений достаточно одного радиационного события, поскольку даже при высоких дозах не отмечено квадратичной компоненты в функции, описывающей кривую «доза — эффект»[94].
4) Хотя удельный вклад конкретных кластерных повреждений несколько отличается для очищенной ДНК и ДНК, облученной в составе клеточного ядра, в целом в суммарном пуле указанных повреждений на один двунитевой разрыв приходится: 1–2 оксипурина: 0,5–0,9 оксипиримидина: 0,75–1,5 потерянных основания. То есть, вклад двунитевых разрывов составляет всего 20–30%.
По крайней мере три типа кластерных повреждений не могли быть детектированы: окисленные пурин + пиримидин; изменения структуры специфического размера и поляризации и две модификации, находящиеся на одной нити без изменений на противоположной (в последнем случае после обработки эндонуклеазами двунитевой разрыв не мог сформироваться).
5) Индукция кластерных повреждений зависит от микроокружения. При облучении ДНК в трис-буфере (обладающем способностью инактивировать свободные радикалы) выход комплексных повреждений составил всего 1–3% от величины, полученной для лучевого воздействия в фосфатном буфере (при отсутствии антиоксидантной активности).
Уровень комплексных повреждений в клетках соответствует таковому при облучении ДНК в трис-буфере. Полагают, что микроокружение ДНК (белки, в частности хроматина, и антиоксиданты) влияет на индукцию и тип кластеров.
6) Выявление кластерных повреждений зависит от методического подхода, практически полностью подавляющего репарацию ДНК: клетки облучали при 0° С и после воздействия помещали в жидкий азот. Методы выделения и хранения ДНК отражались на выходе кластерных повреждений.
7) Базальный уровень комплексных повреждений (без облучения) невелик: в ДНК фага Т7 менее 1 на 106 пар оснований, а в моноцитах [AS57] и в клетках кожи человека [AB13] находится на пределе чувствительности метода, составляя несколько кластерных повреждений на 109 пар оснований (размер эукариотического генома — (5–6) 109 пар нуклеотидов [РЗ14]).

Рисунок 2.5.3. Выход различных типов комплексных повреждений (кластеров) в линии 28SC моноцитов человека в зависимости от дозы рентгеновского облучения; а — двунитевые разрывы, б — оксипуриновые кластеры; в — оксипиримидиновые кластеры; г — кластеры потерянных оснований (на графиках б и в представлены также комплексное повреждение оксипурин (оксипиримидин) + потерянное основание). Малые символы — отдельные опыты; большие светлые символы — M ± m. Значения нормированы авторами к выходу двунитевых разрывов после облучения в дозе 0,5 Гр [AS57].
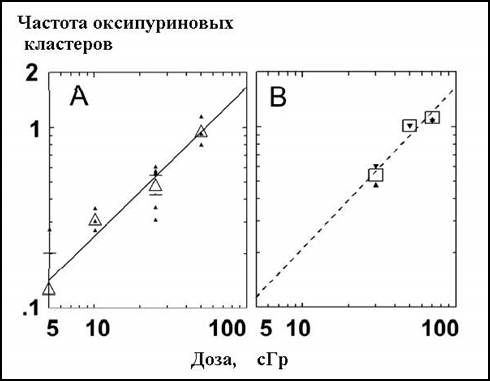
Рисунок 2.5.4. Частота выхода оксипуриновых кластеров при воздействии рентгеновского (А) и плотноионизирующего (B) излучения (ядра 26Fe) на линию 28SC моноцитов человека. Обозначения символов те же, что и на рис. 2.5.3 [AS52].
Б. Сазерленд с соавторами приводили зависимости «доза — эффект» для выхода комплексных повреждений ДНК в облученных клетках и в других работах [AS55]; на рис. 2.5.3 и 2.5.4 мы представили зависимости для наименьших из встретившихся доз, причем редкоионизирующей радиации.
Можно видеть, что выход комплексных повреждений в подвергавшихся воздействию рентгеновского излучения клетках человека почти строго линейно зависит от дозы в диапазоне как малых, так и средних доз. Таким образом, специфические, кластерные повреждения ДНК способны индуцировать даже малые дозы рентгеновского излучения.
Данный вывод подтверждает недавно опубликованное американское исследование [AG5]. Воздействие рентгеновских лучей в дозе 125 мГр на две линии клеток человека MCF-7 (клетки рака груди) и MCF-10A (нераковая линия) приводило к индукции различных типов кластеров окисленных повреждений ДНК. Как и следовало ожидать, более высокий уровень этих повреждений отмечался для раковых клеток по сравнению с нормальными. Но самое удивительное, что кластеры обнаруживались в клетках кожи мышей через 20 недель после облучения в указанной малой дозе in vivo [AG5]. (Нам, правда, не известно, какую линию мышей использовали авторы; возможно, радиочувствительную.)
Итак, кластерные повреждения ДНК могут быть зарегистрированы после облучения in vitro по крайней мере в дозах 50–125 мГр [AG5, AS52, AS57], а in vivo — по крайней мере 125 мГр [AG5]. Но это, вероятно, просто предел чувствительности метода.
На последний вывод нас наводят результаты уже не раз упоминавшегося немецкого исследования К. Роткамма и М. Лёбриха (K. Rothkamm, M. Löbrich) [AR13]). Был использован сверхчувствительный метод регистрации двунитевых разрывов ДНК (по определению числа фокусов гистона γ-H2AX, фосфорилированного в местах их образования). В результате обнаружили, что такие комплексные повреждения ДНК, как двунитевые разрывы, регистрируются в клетках человека уже начиная с сорокакратно меньших, по сравнению с данными Б. Сазерленд с соавторами, доз рентгеновского излучения — 1,2 мГр. А для 10 мГр при регистрации фокусов гистона γ-H2AX имеются данные и других исследователей [AS61].
Опубликована, как уже упоминалось в разделе 1.3, и сверхтонкая модификация метода регистрации однонитевых повреждений ДНК путем кометного анализа, которая, как пишут, «чувствует» до 1–6 мГр на лимфоциты человека [AM3, AS35]. Повторим и здесь, что эта модификация, вероятно, методически сомнительна, поскольку помимо этих работ 1990-х гг. единственной группы авторов [AM3, AS35], других подобных данных нами не обнаружено. Тем не менее, публикации такого рода мы все же имеем.
Поэтому повреждениями ДНК при малых дозах (кластерными и некластерными) нас не удивить, весь вопрос в том, насколько они, эти повреждения, окажутся «жизнеспособными», устояв как перед репарацией, так и перед элиминацией вместе с клетками путем апоптоза. Насколько, так сказать, окажется недостаточным их «процессинг» (в данном случае устранение). А здесь применительно как к кластерам Б. Сазерланд, так и к двунитевым разрывам ДНК К. Роткамма и М. Лёбриха все не так уж и просто (ниже).
* * *
Вернемся к вопросу об уникальности радиационного эффекта по сравнению с другими агентами. Итак, индукция кластерных (комплексных) повреждений — один из главных факторов, обусловливающих специфичность поражающего эффекта радиации. Даже наиболее простые комплексные повреждения — двунитевые разрывы — радиация индуцирует не менее чем в 105 чаще, чем естественный клеточный метаболизм (имеется в виду доля от суммы всех повреждений ДНК) [AF5]. Но следует отличать формирование первичных повреждений ДНК от их конечного выхода, с которым и связаны биологические эффекты (как отмечалось выше, с биологическим эффектом коррелируют не просто уровень повреждений, а нерепарированных повреждений [AD15]). Ведь система защиты — восстановления — элиминации обеспечивает устранение большинства повреждений ДНК, в том числе и радиационно-индуцированных (см. раздел 2.1).
Хотя постулировано, что комплексные повреждения трудно репарируются, но не следует думать, что подобная репарация отсутствует вовсе [AG2, AS57]. Действительно, такие кластерные повреждения, как двунитевые разрывы, успешно воссоединяются (через 1 сут их уровень может снижаться на 90–99% [AG2, AK34, AT5]).
Представленный в [AS57] (см. выше рис. 2.5.3) выход комплексных повреждений авторы нормировали к уровню двунитевых разрывов, индуцированных после облучения в дозе 0,5 Гр. Соответствующая наиболее вероятная величина составляет для этой дозы, по наиболее точным данным (причем совпадающим у разных авторов — см. подборки в [РК14, AU13]), 20 двунитевых разрывов на одну клетку. Согласно [AS57], облучение в минимальной исследованной дозе — 50 мГр — индуцировало различные комплексные повреждения до уровней не более 0,15. Были исследованы три вида кластерных повреждений (помимо двунитевых разрывов), но по крайней мере еще три вида не могли быть детектированы.
Итак, если исходить из [AS57], то всего после облучения в дозе 50 мГр должно индуцироваться 18 (0,15×6×20) кластерных повреждений, не идентичных двунитевым разрывам, в расчете на одну клетку. И — 2 двунитевых разрыва [РК14, AU13]. Порядка 20–30% последних не являются простыми разрывами, они тоже имеют комплексную форму [AN17, AN18].
Продекларированное в [AS57] сохранение линейности кривой выхода кластерных повреждений для области малых доз заставляет полагать, что и ЕРФ способен их индуцировать. ЕРФ составляет 2,38 мГр/год [AU11]. Даже без учета большей ОБЭ a-частиц радона по сравнению с редкоионизирующим излучением получается, что за 48 лет жизни человека ЕРФ (накопленная доза 0,1 Гр) приведет к тому, что, помимо двунитевых разрывов, в каждой клетке организма будет индуцировано 36 комплексных повреждений.
Понятно, что если бы отсутствовали системы защиты от возникновения кластерных повреждений и их репарации, жизнь на Земле была бы невозможна. Относительно систем защиты можно сказать, что они неспособны полностью предотвращать возникновение комплексных повреждений: такие повреждения индуцировались не только в препаратах очищенной ДНК, но и в живых клетках, где защита имеет место. Зато методика приведенных выше экспериментов Б. Сазерленд с соавторами [AG2, AS52, AS55, AS57] по определению уровня комплексных повреждений после радиационного воздействия (облучение при 0°С с последующем помещением клеток в жидкий азот) позволяла сильно подавить репаративные процессы, если не устранить их полностью. Когда же авторы исследовали репарацию кластеров, то клетки облучали при комнатной температуре и, затем, сразу инкубировали при 37°C [AG2].
Таким образом, сложные комплексные повреждения ДНК, отличные от двунитевых разрывов, все же способны репарироваться, как и последние. Это допускают и авторы основополагающих работ [AG2, AS51, AS53, AS54, AS55, AS56, AS57]. В частности, ими продемонстрировано, что при инкубации клеток с селеном (предположительно кофактор репарации) или в клетках с повышенным уровнем фермента репарации белка Fpg эндогенный уровень кластерных повреждений ДНК ниже [AB13].
Принципиальная способность к элиминации даже самых труднорепарируемых комплексных повреждений, представленных в виде особо компактного типа кластеров потерянных оснований, показана в [AG2]. После воздействия γ-излучения в дозах от 1 до 20 Гр на линию 28SC моноцитов человека простые двунитевые разрывы ДНК репарировались в течение 24 ч, как и у других авторов [AK34, AT5]. «Особые» кластеры репарировались гораздо дольше — порядка 14 дней. Но даже при полном отсутствии репарации эти кластерные повреждения могут элиминироваться в процессе репликации ДНК и клеточной пролиферации. Полагают, что данный процесс связан с тем, что при репликации и трансляции родительская ДНК (с кластерным повреждением) расплетается на две дочерних нити с пробелом в одной из них, в результате чего повреждение затем застраивается [AG2].
Здесь надо вспомнить аналогичную картину с репарацией долгоживущих двунитевых разрывов в фибробластах человека после облучения в сверхмалых дозах 1,2 и 5,0 мГр. Это все то же исследование 2003 г. К. Роткамма и М. Лёбриха [AR13]. Хотя для минимальной дозы в 1,2 мГр репарация двунитевых разрывов в покоящихся клетках отсутствовала даже в течение суток, при возобновлении пролиферативного потенциала и клеточного деления все избыточные повреждения ДНК исчезали.
Полагают, что в покоящейся культуре фибробластов репарация индуцируется только при определенном уровне двунитевых разрывов, приблизительно в два раза превышающем спонтанный (по методу регистрации фокусов гистона γ-H2AX — около 0,05 двунитевого разрыва на клетку (0,04–0,06 для 15-ти независимых опытов), т.е., одно спонтанное повреждение на 20 клеток) [95]. При этом дозы в 1,2–20 мГр обеспечивали прибавку в уровне двунитевых разрывов от порядка 0,05 до 0,65 на одну клетку. В среднем как раз и получается 40 двунитевых разрывов на клетку в пересчете на 1 Гр [AR13, AR14], что точно совпадает с данными более ранних исследований, где использовались методики гораздо грубее (см. в [РК14, AU13]).
Существует ряд моментов, касающихся репарации кластерных повреждений, которые можно представить следующими пунктами [AG2, AS53]:
· Каждое повреждение в составе кластерного может репарироваться независимо.
· В процессе репарации кластерные повреждения превращаются в двунитевые разрывы, которые затем воссоединяются обычным образом.
· На репарации кластеров отражаются три фактора: величина пробелов в них, расстояние между комплексными повреждениями и микроокружение (белки), изменяющее конформацию двойной спирали ДНК.
· Близость повреждений в кластерах может приводить к таким конформационным изменениям ДНК, которые затрудняют процесс репарации.
· Клетки могут быть неспособны репарировать некоторые кластерные повреждения, которые накапливаются (особенно в неделящихся клетках, не гибнущих репродуктивно).
· При переходе клеток к репликации ДНК и пролиферации кластерные повреждения (в том числе нерепарированные в начальный период двунитевые разрывы) способны элиминироваться.
Резюмируя сказанное, надо отметить, что если даже некоторые комплексные повреждения не репарируются вовсе, то понятие о малых дозах (см. разделы 1.3 и 1.4) теряет смысл: основываясь на данных [AS52, AS57] можно сказать, что малая доза в 100 мГр γ-излучения приведет к накоплению в клетке ощутимого количества кластерных повреждений: согласно расчетам по [AS57], которые были представлены выше, порядка 40 различных комплексных повреждений на клетку, включая простые и сложные двунитевые разрывы.
Репарация кластерных повреждений ДНК, вероятно, достаточно интенсивный процесс, и именно поэтому в [AG2, AS52, AS55, AS57] пришлось использовать методические подходы, ее устраняющие. Можно полагать, что в противном случае просто не удалось бы обнаружить комплексные повреждения ДНК в клетках после облучения в дозах менее 0,1–0,2 Гр, поскольку они были бы репарированы.
К этому. Сторонник гормезиса [AK23], основатель учения об апоптозе [AK21] Сохеи Кондо (Sohei Kondo)[96] в своем комментарии на Сообщение NCRP-136 [AN8] отмечал, что маловероятно выдвинутое там предположение о специфичности радиационно-индуцированных двунитевых разрывов по сравнению со спонтанными, которая, де, реализуется у первых в структуре, значительно затрудняющей репарацию [AK22].
С. Кондо приводит пример с редким генетическим аутосомным заболеванием — синдромом поломок Неймегена, который связан с дефектами в репарации двунитевых разрывов ДНК. Больные этим синдромом в 40% случаев заболевают к 21 году лимфомами, лейкозами или другими опухолями, а врожденные патологии характерны для 50% пациентов [AN16].
Ясно, что высокая частота индукции опухолей и врожденных уродств обусловлена дефектами в репарации именно спонтанных двунитевых разрывов ДНК. Отсюда следует, что в отсутствии репарации и спонтанные двунитевые разрывы ДНК столь же гибельны для клетки, как и индуцированные генотоксическими агентами. Таким образом, заключает С. Кондо, это никак не совмещается с положением о специфических повреждающих особенностях радиационно-индуцированных двунитевых разрывов [AK22].
Можно добавить, что мутантные клетки, у которых отсутствует или снижена репарация двунитевых разрывов, проявляют нестабильность генома. Таких примеров много; приводим два [AD14, AK11].
Из материалов данного подраздела мы можем сделать вывод, что, хотя для действия ионизирующего излучения на клетку и характерна значительная специфичность повреждающего эффекта по сравнению со спонтанными нарушениями/модификациями структуры ДНК, эта специфичность не распространяется настолько далеко, чтобы свидетельствовать о накоплении нерепарированных повреждений при любых дозах радиационного воздействия. Даже присущие радиации комплексные повреждения ДНК, простые и сложные двунитевые разрывы, должны в большинстве своем успешно устраняться в условиях жизни на Земле, когда воздействия ЕРФ невозможно избежать. Следует помнить, что утверждение об отсутствии репарации комплексных повреждений ДНК является только теоретическим постулатом [AG6, AW2, AW3, AW4]. Реальность свидетельствует, как мы видели выше, об обратном. В то же время, хотя подобные повреждения и репарируются, но, вероятно, «в большинстве своем», т.е. не все. И именно оставшиеся нерепарированными или неверно репарированные комплексные двунитевые разрывы и другие сложные повреждения ДНК отвечают за мутагенез [AB10]. Именно эти события могут вызывать значительные по размеру делеции и пробелы в структуре генома, приводящие к канцерогенным мутациям. Вот только остается вопрос о дозах, которые индуцируют столько труднорепарируемых повреждений, чтобы после их процессинга что-то все же оставалось бы.
Дата добавления: 2015-09-10; просмотров: 96 | Поможем написать вашу работу | Нарушение авторских прав |